




Водно-болотное угодье бассейна оз. Ханка, фото В. Герштейн
оз. Кривое, фото Д. Красников
Колония чаек на о. Сосновый, фото Ю. Сушицкий
Разнотравные луга, фото В. Герштейн



Телефон: +7 (42352) 20887
e-mail: priroda24@yandex.ru
|
|









![]()
![]()

![]()
В этом выпуске мы будем знакомить читателей с видовым разнообразием рыб, редко встречающихся в бассейне оз. Ханка.
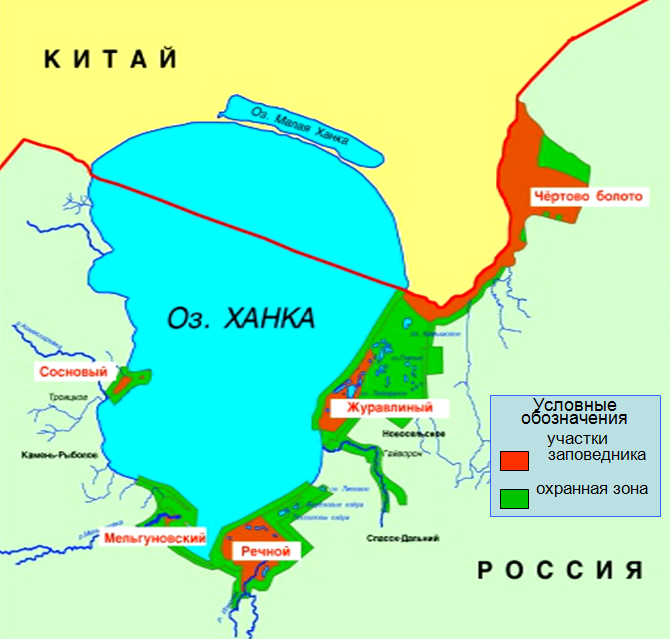
И первый представитель данной категории ихтиофауны, о которым мы хотим поведать - Пескарь-лень (Sarcocheilichthys sinensis Bleeker, 1871).
Пескарь-лень
(Sarcocheilichthys sinensis Bleeker, 1871)
Класс Костные рыбы – Osteichthyes
Отряд Карпообразные – Cypriniformes
Семейство Карповые - Cyprinidae
 |
Пресноводный вид. Промыслового значения не имеет. В отдельных частях ареала редок. В Приморье встречается в бассейнах оз. Ханка и р. Уссури. Держится преимущественно в низовьях рек, в русле и протоках. В бассейне оз. Ханка отмечены поимки пескаря в устьевых зонах р. Спасовка и р. Илистая. Общий ареал охватывает водоёмы Восточной Азии от бассейна р. Амур до рек Кореи.
Пескарь-лень достигает более крупных размеров, чем другие пескари – длины до 30 см и массы 300 г. Продолжительность жизни 8-9 лет.
Тело овально-удлиненное, сжатое с боков. Окраска тела молоди золотистая, с 4-я поперечными полосками. У взрослых полосы малозаметны, тело тёмное с лиловатым оттенком. Во время нереста у самцов на голове появляются эпителиальные бугорки, у самок – очень короткий яйцеклад. Рот нижний, усиков нет. Рыло тупое, закруглённое. Часть нижней челюсти, не покрытая губами, имеет вид совочка с заострённым краем, покрытым роговым чехликом.
Половозрелым становится на 3-м году жизни при длине более 10 см. Пескарь-лень порционно мечет очень крупную пелагическую икру, имеющую матовую оболочку, однако часто откладывает её в мантийную полость пресноводных моллюсков (беззубок, ножичков). Нерест происходит с начала июня до второй половины июля при температуре воды выше 240С. Плодовитость колеблется от 5 до 18 тыс. икринок. Развитие продолжается около 4 суток, выклев при длине 7,5 мм. Выклюнувшиеся личинки ведут пелагический образ жизни. Питается донными беспозвоночными (личинки хирономид, подёнок, ручейников), соскабливая их с подводных предметов своей приостренной нижней челюстью.
Литература:
- Бушуев В.П. Пресноводные и эстуарные рыбы Приморья : справ. / В.П. Бушуев, Е.И. Барабанщиков. – Владивосток : Дальрыбвтуз, 2012. – 314 с.
Материал подготовлен младшим научным сотрудником
ФГБУ «Государственный заповедник «Ханкайский»
Бруневской Е.Ю.
Редкие рыбы озера Ханка
Несмотря на громадную площадь водного бассейна озера Ханка, разнообразие рыб и их обилие – всё это не может служить поручительством, что рыбные богатства останутся навсегда. Напротив, путём хищнических способов ловли можно быстро истребить природные богатства.
Одной из задач, стоящих перед заповедником «Ханкайский», является сохранение популяций редких видов животных, в том числе и рыб, обитающих в бассейне озера Ханка. В озере встречаются 9 видов рыб, включенных в Красные книги различного статуса: амурский осётр, калуга, чёрный амур, чёрный амурский лещ, сом Солдатова, желтощёк, мелкочешуйный желтопёр, ауха (китайский окунь) и косатка-крошка.
Калуга – Huso dauricus (Georgi, 1775).
 |
Является эндемиком амурского бассейна. В Красный список МСОП занесена зейско-буреинская популяция в статусе первой категории как «находящаяся под угрозой исчезновения популяция эндемичного вида». Одна из крупнейших пресноводных рыб, достигает длины 5,6 м и веса 1140 кг, живет более 55 лет. Половая зрелость у самок наступает не ранее 16-17 лет, по достижении длины более 2 м. Самцы созревают на один-два года раньше самок.
Типично русловая рыба. Размножается калуга на галечниково-песчаных грядах в русле Амура на глубине от 3 до 7 метров. Сроки нереста май-июнь. Совершает протяженные нагульные и нерестовые миграции на тысячу и более километров. На акватории заповедника, как и в озере Ханка в целом, всегда была редка.
Амурский осётр – Acipenser schrenckii (Brandt, 1869).
 |
Является эндемичным пресноводным промысловым видом бассейна. В Красный список МСОП занесена зейско-буреинская популяция в статусе первой категории как «находящаяся под угрозой исчезновения популяция эндемичного вида». Типично русловая рыба и в озёра для нагула заходит очень редко. Достигает длины более трёх метров и массы более 250 кг приблизительно за 65 лет жизни. Самка впервые созревает и нерестится в возрасте 10-14 лет, по достижению длины тела более 105 см и массы 6-12 кг. Самцы становятся половозрелыми в 7-12 лет, достигнув длины тела 95-110 см и массы 4-10 кг. Нерест амурского осётра проходит с конца мая до середины июля при температуре воды 120С-180С. Нерестилища расположены на галечниковых участках реки Амур с быстрым течением и глубиной 3-6 м. В настоящее время на акватории заповедника, как и в озере Ханка в целом, очень редок.
Китайский окунь (ауха) – Sinipercachuatsi (Basilewsky, 1855).
 |
Пресноводный жилой вид. Включён в Красную книгу РФ по рекомендации М.Л. Крыхтина в статусе «вторая категория», как сокращающийся в численности вид, а также в Красную книгу Приморского края. Крупный пелагический хищник, достигает длины 70 см и массы 9 кг. Летом держится как в русле, так и в пойменных водоёмах. Половозрелым становится на пятом году жизни при длине 32-34 см. Нерест порционный в июне-июле при температуре воды 200С -260С. В последние годы отмечается значительный рост численности аухи. В настоящее время на акватории заповедника, как и в озере Ханка в целом, обычен.
Мелкочешуйный желтопёр -Plagiognathops (Xenocypris) microlepis (Bleeker, 1871).
 |
Один из самых теплолюбивых видов рыб амурского комплекса. Был включён в Красную книгу РФ по рекомендации М.Л. Крыхтина в статусе «первая категория» как вид, находящийся под угрозой исчезновения, а также в Красную книгу Приморского края. Населяет крупные озёра и равнинные реки. Где предпочитает зарастающее водной растительностью прибрежье. Половой зрелости достигает на 4-5 году жизни при длине около 35 см и массе 700-800 г. Нерест в конце июня - начале июля. Продолжительность жизни более шести лет. В настоящее время на акватории заповедника и в озере Ханка редок.
Сом Солдатова - Silurus soldatovi (G. Nikolsky et Soin, 1948).
 |
 |
Был включён в Красную книгу РФ по рекомендации Н.С. Пробатова и М.Л. Крыхтина в статусе «первая категория» как вид, находящийся под угрозой исчезновения. В настоящее время этот вид имеет статус второй категории, как сокращающийся в численности узкоареальный вид. Также внесён в Красную книгу Приморского края. Относительно теплолюбивая рыба, поэтому избегает горные и предгорные притоки. Самки созревают в возрасте 4-6 лет при длине 90-100 см и массе 8-10 кг, а самцы – в возрасте 4-5 лет при длине 85-90 см и массе 6-7 кг. Продолжительность жизни более 20 лет. Растёт очень быстро. Достигает длины 2 м и массы 40 кг. Нерест проходит в июне, икру откладывая на растительность.В настоящее время на акватории заповедника и в озере Ханка редок.
Чёрный амурский лещ - Megalobramamantschuricus (Basilewsky, 1855).
 |
В Красной книге РФ 2001 г. имеет название Megalobramaterminalis.
Включён в Красную книгу РФ и в Красную книгу Приморского края. Пресноводный жилой вид. Обитает как на озёрных участках, так и в приустьевых частях рек. Протяжённых миграций не совершает. Половозрелым становится на 6-м году жизни при длине около 30 см. Нерестится в весенне-летний период. В настоящее время на акватории заповедника и его охранной зоне обычен.
Чёрный амур - Mylopharyngodon piceus (Richardson, 1846).
 |
Вид включён в Красную книгу РФ по рекомендации М.Л. Крыхтина в статусе «первая категория» как вид, находящийся под угрозой исчезновения и Красную книгу Приморского края. Достигает длины более 100 см и массы 35 кг. Продолжительность жизни более 13 лет. Приурочен к слабопроточным участкам, вблизи скоплений моллюсков. Половой зрелости достигает в возрасте 7-10 лет при длине 66-80 см. В озере Ханка не размножается. В настоящее время на акватории заповедника и в озере Ханка редок.
Желтощёк - Elopichthysbambusa (Richardson, 1845).
 |
Включён в Красную книгу РФ в статусе «первая категория» как вид, находящийся под угрозой исчезновения. Крупная рыба, достигающая длины 170-200 см и массы 30-40 кг. Половой зрелости достигает на шестом году жизни при длине не менее 60 см. В озере Ханка не размножается, заходит только для нагула и зимовки из р. Уссури. В настоящее время на акватории заповедника и в озере Ханка малочислен.
Косатка-крошка - Pelteobagrus mica (Gromov, 1970).
 |
Ошибочно внесена в Красную книгу Приморского края. Мелкая рыба длиной 3-5 см. Обычные места обитания косатки-крошки – прибрежные участки русла рек со слабым течением и песчано-илистым грунтом. Отмечена и в пойменных озерах. Довольно многочисленна в озере Ханка и во всей придаточной системе.
10 июля – День действий против рыбной ловли
На основании Правил рыболовства для Дальневосточного рыбохозяйственного бассейна ежегодно с 20 апреля по 20 июля в озере Ханка, в устьях впадающих в него рек и на расстоянии 1 км от устья вверх по течению и в разливах запрещается любительское и спортивное рыболовство всех видов водных биоресурсов (за исключением рыболовства удебными орудиями добычи (вылова) всех видов и наименований. Данный период времени объявлен трехмесячником по охране нерестующих видов рыб для поддержания численности и видового разнообразия рыбных ресурсов амурского комплекса, в том числе видов, занесённых в списки Красных книг различного статуса.
В иных случаях рыбалка разрешена везде в определенные сроки, за исключением территории заповедников, частных владений и предприятий по разведению рыб.
Согласно Правил рыболовства для Дальневосточного рыбохозяйственного бассейна № 385 от 21 октября 2013 года, с изменениями на 28 октября 2015 года (редакция, действующая с 1 января 2016 года), «…граждане вправе осуществлять любительское и спортивное рыболовство на водных объектах рыбохозяйственного значения общего пользования свободно и бесплатно в соответствии с Правилами рыболовства».
При любительском и спортивном рыболовстве запрещается:
- применение аханов (сетей с размером (шагом) ячеи 90 мм и более), самоловов, донных и пелагических тралов, «фонарей», капканов, острог;
- установка в водных объектах рыбохозяйственного значения заколов и других видов заграждений;
- прекращать доступ кислорода и воды в водный объект рыбохозяйственного значения посредством уничтожения источников его водоснабжения, а также осуществлять спуск водных объектов рыбохозяйственного значения с целью добычи (вылова) водных биоресурсов (за исключением прудов для товарного рыбоводства, находящихся вне русел естественных водотоков и оборудованных гидротехническими сооружениями, регулирующими подачу и сброс воды);
- установка крючковых орудий добычи (вылова) с количеством блесен или крючков, превышающим установленное Правилами рыболовства.
При любительском и спортивном рыболовстве на озере Ханка без путёвок запрещается применение драг, ставных, плавных и иных видов сетей, неводов, бредней, вентерей (верш), мереж (рюж), ручных сачков, подъемных сеток, петель, захватов, фитилей.
За нарушение закона о пользовании водными биоресурсами предусмотрены следующие наказания и штрафы:
1. За нарушение правил вылова рыбы и иных правил об осуществлении рыболовства (КоАП ст.8.37 ч.2).
2. За ловлю рыбы во время нереста.
3. За незаконный вылов рыбы посредством использования запрещённых орудий лова, с причинением ущерба в крупных размерах, во время (и в местах) нереста, в запрещённых местах для лова (ст.256 УК РФ). Наказание: штраф в 100000-300000 руб. для граждан, либо арест на 6 мес.
4. За уничтожение/повреждение специальных знаков, которые определяют границы особо охраняемых объектов, охраняемых зон водных объектов и пр. Наказание: штраф, равный 300-500 руб. для граждан.
5. За рыбалку без лицензии (при ее необходимости)
6. За уничтожение редких видов рыб (из Красной книги).
Ежегодно во второе воскресенье июля любой гражданин нашей страны может проявить свою социальную активность и поучаствовать в общественной экологической акции против безмерной рыбной ловли. Для этого достаточно немного "укротить" свой азарт заядлого рыболова. На смену этому, далеко не безобидному хобби, было бы неплохо провести агитацию среди коллег и товарищей по подобному "ремеслу" не превращать любительское рыболовство в варварское истребление рыбных запасов. Важно чаще беседовать с детьми и подростками, объяснять им, что следует беречь рыбные ресурсы страны. Кроме того, можно завести дома аквариум с многочисленными и экзотическими рыбками – пусть это будет ваш символический протест против рыбной ловли вне закона!
Скачайте универсальный справочник по редким рыбам озера Ханка!
Ниже размещена ссылка на скачивание справочника по рыбам озера Ханка, занесенным в Красные книги РФ и Приморского края (справочник подходит к использованию на мобильных устройствах).
 |
Материал подготовлен младшим научным сотрудником
ФГБУ «Государственный заповедник «Ханкайский»
Бруневской Е. Ю.
совместно со специалистами отдела экологического просвещения
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
После зимы, с наступлением теплых дней быстро пробуждаются и рано зацветают некоторые многолетники-первоцветы. Цветут они обычно уже в апреле (в отдельные теплые годы - с конца марта) до середины мая. Таким образом, все они относятся к ранневесенней флоре. Первые цветы имеют свою прелесть. Может потому, что им особо радуешься, они самые долгожданные. Их немного, и потому разглядываешь каждый цветочек.
В природе ничто не происходит "просто так". Если что-то имеет в ней место, значит, этому есть свои причины. Вернемся к нашим первоцветам и зададим себе вопрос: "зачем они цветут так рано весной?". Иными словами, "в чем биологический смысл данного явления (ранневесеннего цветения) этой группы растений?".
Для начала вспомним, что растениям для нормальной жизнедеятельности необходим солнечный свет. Именно на свету в зеленых органах растений происходят процессы фотосинтеза, когда из неорганических веществ (воды и углекислого газа) образуются органические вещества - углеводы, которые потом растения используют для своего развития. Таким образом, достаточное количество солнечного света является необходимым условием нормального развития растений. Вся жизнь растений проходит в постоянной борьбе за свет.
Деревья и кустарники еще не оделись листвой, ничто не мешает солнечному свету беспрепятственно проникать до самой земли. Именно это обстоятельство и является основной причиной того, что многие виды растений в процессе эволюции "выбрали" для своего цветения это время года. Кроме того, земля после схода снега насыщена влагой, которая также является необходимым условием для нормального развития растений. В это время года, однако, еще довольно прохладно, и раннецветущим растениям пришлось приспосабливаться к этому фактору. Эти растения отнесли в особую группу эфемероидов, светолюбивых многолетников с коротким вегетационным периодом и длительным покоем. Эфемероиды представляют собой яркий пример адаптации травянистых растений листопадных лесов к влажному и светлому весеннему периоду (до распускания почек на деревьях). Они отличаются необычайной "торопливостью"- появляются на свет тотчас же после схода снега и быстро развиваются, несмотря на весеннюю прохладу. Через неделю-другую после появления на свет они уже цветут, а еще через две-три недели у них появляются плоды с семенами. Сами растения при этом желтеют и полегают на землю, а затем надземная их часть отмирает. Происходит все это в самом начале лета, когда, казалось бы, условия для жизни лесных растений самые благоприятные - достаточно тепла и влаги. Период глубокого затенения эфемероиды переживают в состоянии летнего покоя, в виде подземных органов - луковиц, клубней, корневищ. Несмотря на краткие сроки вегетации, эфемероиды играют существенную роль в создании биологической массы травяного покрова и в общем круговороте веществ в листопадных лесах. Эфемероиды после отмирания и разложения надземной части вносят в почву значительное количество питательных веществ (особенно калия и азота).
| Название | Начало цветения | Массовое цветение | Конец цветения | Плодоношение |
| Адонис амурский | вторая половина марта | 5 – 20 апреля | начало мая | конец апреля – середина июня |
| Гусиный лук Террачино |
середина апреля | начало мая | середина мая | май, июнь |
| Калужница болотная | 2-я декада апреля | начало мая | конец мая | |
| Ветровник вильчатый | 3-я декада апреля | начало мая | 2-я декада мая | |
| Лапчатка земляничная | с 15 – 20 апреля | с 28 апреля – 10 мая | начало июня | с середины мая до конца июня |
| Прострел даурский | с середины 3-й декады апреля | начало мая | середина мая | май - июль |
| Сердечник трехнадрезанный | вторая декада апреля | конец апреля - начало мая | середина мая | май, июнь |
| Фиалка холмовая | начало апреля | 2-я декада апреля – начало мая | 2-я декада мая | со 2-й декады мая |
Литература:
- Горышина, Т. К. Ранневесенние эфемероиды лесостепных дубрав (исследования по экологии, физиологиии фитоценологии). - Л., 1969;
- Смирнова, О. В. Структура травяного покрова широколиственных лесов. - М., 1987.
Материал подготовлен науч. сотрудником
ФГБУ «Государственный заповедник «Ханкайский»
Мазурок Н.Н.
Наука - людям
ЛЕБЕДИ
Лебеди (лат. Cygnus) - род птиц из отряда гусеобразных семейства утиных.
Лебеди - очень крупные птицы с белым оперением и с длинной тонкой шеей. Лапы у взрослых черные, довольно короткие, из-за чего лебеди, передвигаясь по земле, производят несколько неуклюжее впечатление. С воды взлетают с разбега. Нырять не могут, кормятся, погружая в воду шею или переднюю часть тела. Самок и самцов внешне весьма трудно различить. Лебедей от гусей отличает более длинная шея, позволяющая в более глубоких водах обыскивать дно в поисках пищи, а также их размер, по которому они являются самыми крупными водными птицами.
Обитатели озер, рек, берегов морей. Потомство выращивается обоими родителями. Питаются водной растительностью. Лебеди очень рано прилетают с зимовок и поздно улетают на юг.
На территории, вошедшей в заповедник «Ханкайский», и Приханкайской низменности зарегистрированы встречи трех видов лебедей: лебедь-шипун, лебедь-кликун, малый лебедь. Все три вида включены в Красную книгу Приморского края, а малый лебедь и в Красную книгу России. В настоящее время на рассматриваемой территории гнездится только лебедь-кликун, малый лебедь встречается на пролете, а лебедь-шипун должен считаться исчезнувшим видом с крайне редкими эпизодическими залетами.
Лебедь-шипун – Cygnus olor (Gmelin, 1789)
 |
Рисунок из «A field guide to the BIRDS OF KOREA»
Самый крупный из лебедей. Размах крыльев лебедя-шипуна около 240 см. Иногда на голове и шее заметен рыжеватый налет. Клюв красный с черным кончиком и основанием, на лбу черный шишковидный нарост, хвост слегка клиновидный. У молодых клюв розовый с черным кончиком и основанием, нароста на клюве нет. Брюхо буроватое, ноги красные. У сидящей на воде птицы шея (более толстая, чем у других видов лебедей) изогнута в виде буквы S, а крылья приподняты. При раздражении издает характерный шипящий звук, по которому и получил свое название.
Гнездится в тростниковых зарослях на озерах и в дельтах рек северо-запада и юга России и Дальнего Востока. Гнездо из стеблей тростника строит на заросших водоемах. В кладке 6-8 голубоватых яиц. Зимует на побережьях морей.
В прошлом гнездился как на Ханке, так и на озёрах крайнего юго-запада Приморья. Относительно его гнездования в ханкайском бассейне в литературе имеются некоторые сомнения, касающиеся достоверности данных, полученных Н.М. Пржевальским, который не добывал, а лишь наблюдал птиц с большого расстояния, и не был точно уверен в правильности их видового определения. Тем не менее, в пользу обитания здесь лебедя-шипуна в прошлом свидетельствуют археологические находки костей, обнаруженных при исследованиях расположенного у границ заповедника археологического памятника эпох неолита и бронзы «Синий Гай».
Единственный случай залёта шипуна на Приханкайскую низменность в ХХ столетии зарегистрирован в начале апреля 1966 г., когда одна особь наблюдалась в окрестностях с. Гайворон.
Лебедь-кликун – Cygnus cygnus (Linnaeus, 1758)
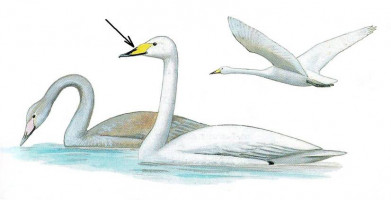 |
Рисунок из «A field guide to the BIRDS OF KOREA»
У лебедя-кликуна клюв желтый с черным кончиком, причем желтый цвет заходит вперед за ноздри. У молодых голова и шея буроватые, брюхо белое, клюв и ноги розоватые. Плавает обычно держа шею вертикально.
Гнездится отдельными парами на зарастающих озерах от тундры до лесостепи. Гнездо – крупная постройка из мха, травы или тростника на мелководье у берега или на островке. В кладке 4-6 белых или желтоватых яиц. Зимует на побережьях морей, иногда на незамерзающих озерах юга России. Голос – громкое трубное «ганг-го….ганг-го».
История гнездящейся популяции лебедя-кликуна на Ханке весьма интересна. Как и шипун, данный вид размножался на Ханке во второй половине 19-го века, однако впоследствии тоже исчез. В начале 60-х годов 20-го века он вновь был встречен здесь в летнее время, и высказывалось мнение о его гнездовании, хотя достоверно гнёзда обнаружены не были. Лишь в конце 70-х годов прошлого столетия были найдены первые гнёзда, причём все они были устроены однотипно: яйца откладывались на утрамбованные лебедями хатки ондатры. В полной кладке содержится от 4 до 6 яиц. Яйца имеют размеры 109,8-124,3 х 71,4-77,3 мм. В выводках наблюдалось от 2 до 6 птенцов.
Численность гнездящейся популяции кликуна здесь (включая долину р. Сунгача) в конце прошлого века была очень невелика, составляя от 1 до 6 гнездящихся пар, в дополнении к которым проводит лето, не приступая к размножению до 30 неполовозрелых особей. Большинство гнездящихся и летующих птиц придерживались бассейна р. Гнилая и прилежащих к нему озёр, вошедших в охранную зону заповедника. До конца прошлого столетия кликуны гнездились исключительно на изолированных озёрах, куда нет доступа моторным лодкам. Однако, в 2002 г. на Гнилых озёрах, связанных с Ханкой посредством р. Гнилая, было обнаружено 4 гнездящиеся пары. И, хотя выявить общую численность гнездящихся на низменности лебедей не удалось, она наверняка составила не менее 10 пар. Таким образом, вновь возникшая местная популяция лебедя-кликуна проявляет некоторую тенденцию роста, явно связанную со снижением фактора беспокойства на потенциальных местах размножения, что в первую очередь обусловлено созданием Ханкайского заповедника.
Интенсивность как весеннего, так и осеннего пролётов подвержена весьма значительным межгодовым изменениям. Обычно видимый пролёт выражен крайне слабо, но в некоторые годы кликун вполне обычен, причём наблюдаются скопления, насчитывающие порой многие сотни особей. Любопытно отметить, что с середины 80-х до начала 90-х годов 20-го века численность пролётных и останавливающихся весной на Ханке лебедей заметно возрастала, и птицы стали охотно кормиться на рисовых полях. К сожалению, такая ситуация сохранялась очень недолго и в самом конце прошлого столетия картина резко изменилась в худшую сторону, поскольку зона рисосеяния в пределах Приханкайской низменности в этот период резко сократилась, а на оставшихся полях обычно велась массированная охота.
Наиболее ранние встречи зарегистрированы 2 марта, пролёт длится до конца апреля, хотя значительное число птиц (до сотни особей), может держаться на озёрах низменности (в частности в бассейне р. Гнилая) в течение всего мая и даже июня. Осенний пролёт протекает в октябре и начале ноября, а наиболее поздние встречи зафиксированы 18 ноября.
Малый лебедь – Cygnus bewickii Yarrel, 1830
 |
Рисунок из «A field guide to the BIRDS OF KOREA»
Малый лебедь похож на лебедя-кликуна, но заметно меньше, шея и клюв короче, желтый цвет на основании клюва не доходит до ноздрей, а передний край желтого пятна обычно почти вертикальный. У молодых птиц кончик клюва темно-серый.
Гнездится на озерах в низменных тундрах и заболоченной лесотундре к востоку от полуострова Канин. Гнездо из мха и осок на кочке или сухом островке у берега. В кладке 3-4 желтоватых яйца. Голос слабее и звонче чем у клоктуна.
На Ханке малочисленный пролётный вид. Встречается главным образом в апреле и октябре, часто совместно с предыдущим видом, однако в численности значительно уступает ему, составляя в разные годы от 6 до 16,9% от общего числа учтённых лебедей (Глущенко, Бочарников, Шибнев, 1995). Отдельные особи наблюдались в не типичное для пролёта время, так одна из них была зарегистрирована на рисовых полях у с. Сосновка 30 мая 1993 г..
Использованная литература
Бёме Р.Л., Динец В.Л., Флинт В.Е., Черенков А.Е. Птицы. Энциклопедия природы России. М.: 1998. С. 70-73.
Гладков Н.А. и др. Определитель птиц СССР. Ярославль. 1964. С. 175-177.
Красная книга Приморского края. Животные. 2005. Редкие и находящиеся под угрозой исчезновения виды животных. Официальное издание. Владивосток, АВК «Апельсин». С. 220-222.
Позвоночные животные заповедника «Ханкайский» и Приханкайской низменности: Монография. – Владивосток: ООО РИЦ «Идея», 2006. С. 101-103.
Материал подготовлен старшим науч. сотрудником
ФГБУ «Государственный заповедник «Ханкайский»
Герштейном Владимиром Витальевичем
Наука - людям
14 сентября - Всемирный день журавля
В заповеднике «Ханкайский» и на Приханкайской низменности гнездятся два вида журавлей: японский и даурский. Оба эти вида внесены во всевозможные Красные книги и списки (Приморского, Хабаровского краев, Дальнего Востока, Российской Федерации, Японии, Южной Кореи, Китая и Азии, МСОП) как виды, находящиеся под угрозой исчезновения, спасение которых невозможно без осуществления специальных мер.
 |
 |
Фото 1, 2 - японский журавль
 |
Фото 3. Даурский журавль
Также встречается в нашем регионе в период пролёта чёрный журавль, и очень редко, в форме залётов отмечаются стерх, красавка и серый журавль (отмечен только в китайском секторе Приханкайской низменности).
В период размножения журавли связаны с заболоченными местностями. Семьи сохраняются в течение года. Полового деморфизма в окраске нет, по размерам самцы обычно несколько крупнее. В период размножения журавли строго территориальны и держатся парами, во время миграций и на зимовках ведут групповой и стайный образ жизни. Питаются как растительной, так и животной пищей. Тип развития птенцов - выводковый.
В полёте ноги и шея вытянуты; на земле в спокойном состоянии птицы кажутся сгорбленными, при тревоге выпрямляются и вытягивают шею вверх. Стая в полёте обычно выстраивается в форме клина.
У японских журавлей предпочтительным биотопом являются обширные вейниково-осоковые и осоково-пушицевые болота. Такие биотопы располагаются в понижении микрорельефа местности. Японский журавль для гнездования использует малодоступные центральные части болотных массивов. В отличие от японских, даурские журавли обитают на слабо увлажнённых пространствах вейниково-осоковых болот и осоково-разнотравных лугов, на возвышениях микрорельефа. Индивидуальные территории даурских журавлей часто сочетают естественную среду обитания, с разной степенью нарушенную человеком, и сельхозугодья.
Японский журавль (Grus japonensis P.L.S. Muller,1776) - самый крупный из наших журавлей, с размахом крыльев около 2,7 м, высотой около 1,6 м, массой до 11 кг. Продолжительность жизни японских журавлей в природе, по данным кольцевания, составляет 13 лет.
В местах гнездования этот журавль очень осторожен, особенно у гнезда, держится скрытно, человека не подпускает ближе полукилометра и сравнительно редко попадается на глаза. На зимовках эти птицы значительно более доверчивы.
От стерха, в окраске которого тоже доминирует белый цвет, японский журавль легко отличается белыми концами крыльев, чёрными шеей и задней стороной крыльев, от других наших журавлей - белой окраской тела, от дальневосточного аиста - белыми концами крыльев, чёрными ногами. На темени у японского журавля присутствует лишённый оперения участок кожи красного цвета.
Как и у других журавлей, поведение японского журавля в значительной степени ритуализировано, причём определённые ситуации сопровождаются не только соответствующими позами и движениями (см. рис. 4), но и голосовыми сигналами. Голос японского журавля очень громкий и сильный, особенно во время унисонального дуэта. Журавли исполняют его стоя на расстоянии 1-3 м друг от друга, повернувшись в одну сторону или друг к другу; обе птицы кричат одновременно, причём крик самца несколько опережает крик самки, таким образом получается единое полифоническое звучания. Птицы во время унисонального дуэта быстро вытягивают шеи, направляя клювы вверх и вперёд, самец поднимает над спиной сложенные крылья. Крик в унисон отмечен в утренние часы, при смене родителей на гнезде, после вылупления одного из птенцов, при опасности. Унисональный дуэт исполняется и вне сезона размножения: на зимовках, в местах кормёжки, особенно интенсивно - в конце зимы и начале весны.
Отличительная особенность прилёта японских журавлей - прилёт парами и семейными группами в сжатые сроки. Во время осеннего и весеннего пролёта в скоплениях японских журавлей можно наблюдать хорошо известные «танцы»: несколько птиц одновременно демонстрируют ряд особых движений - высокие прыжки с развёрнутыми крыльями, поклоны и т. п. В «танцах» принимают участие как взрослые, так и молодые птицы. Обычно «танцы» наблюдаются после кормёжки, когда одна из птиц начинает подпрыгивать, развернув крылья, и увлекает других, вскоре оживление переходит в коллективный «танец», захватывающий всю стаю. «Танцы» японских журавлей, бесспорно, полифункциональны, они отражают состояние птиц и служат выходом энергии, играют определённую роль в образовании супружеских пар и поддержании связей в группе.
Основной гнездовой ареал японского журавля охватывает Приморье, Приамурье, Северо-Восточный Китай, в Японии изолированная популяция обитает на о-ве Хоккайдо. Зимовки расположены на Корейском полуострове и в Китае. Основные места обитания в гнездовой период - обширные кочкарниковые осоково-пушицевые и осоково-моховые болота и осоково-вейниковые заболоченные луга, с хорошим обзором, располагающиеся обычно по долинам рек и в котловинах озёр. Они имеют сравнительно твердую, нетопкую почву и весной ежегодно заливаются водой, а к середине лета в значительной степени пересыхают.
Для устройства гнезда птицы всегда выбирают участки, покрытые невыгоревшей высокой сухой осокой или вейником с примесью тростника и залитые водой. Гнёзда располагаются на кочках или плоской дерновине, возвышаясь над водой на 15-20 см, и представляют собой плоскую овальную платформу.
Половая зрелость у журавлей наступает в возрасте 3-4 года, они моногамны, часто пары сохраняются на протяжении всей жизни. Японский журавль прилетает на места гнездования во второй половине марта, когда земля ещё не оттаяла, а возвраты холодов и метелей представляют закономерное явление. Гнездовые участки журавли занимают в апреле; размер индивидуального участка составляет 6 - 12 км2, расстояние между соседними гнёздами 3 - 4 км. Кладка состоит из двух яиц, реже из одного. В насиживании принимают участие оба партнёра. Ночью на гнезде сидит, как правило, самка, самец проводит ночь поблизости от гнезда, но в течение дня он сменяет её 2-3 раза. К концу инкубационного периода роль самца в насиживании сокращается. Вылупление птенцов происходит, в основном, во второй половине мая - первой декаде июня. Первый птенец проводит в гнезде около двух суток, второй покидает его сразу после того, как обсохнет. Агрессивного гнездового поведения у птенцов японского журавля не отмечено, обычно жизнеспособны оба птенца. Пуховиков, покинувших гнездо, в первые дни жизни взрослые обогревают, присаживаясь на сухих местах. Пока одна из птиц согревает птенцов, другая собирает корм поблизости. Позже, когда птенцы окрепнут, один из журавлей держится с ними, а другой ищет корм в 20-30 м, а поймав добычу, кормит пуховиков, тогда как другой партнёр в свою очередь удаляется на поиски корма. Если поблизости корма достаточно, выводки держатся на кормовом участке 1-2 недели. При беспокойстве родители уводят птенцов в наиболее глухие участки болота; при пересыхании болот выводки перемещаются в более сырые места.
Кормовые биотопы используются в течение всего гнездового сезона, с учётом представленного в них пищевого разнообразия, а также доступности объектов питания. В гнездовой период у партнёров, чаще всего это самцы, кормовые стации могут быть на расстоянии двух и более километров от гнезда и не входить в индивидуальный участок. Смена кормовых биотопов обуславливается не столько истощением кормовых ресурсов, сколько появлением других объектов питания в большем количестве и доступных в добывании, при этом пребывание журавлей в увлажнённых биотопах летом составляет 70% встреч. В питании японский журавль использует широкий спектр кормов, соотношение растительной и животной пищи в различные периоды меняется, однако преобладает животная пища, только на зимовках и в определённые периоды он переходит в значительной мере на растительные корма. В качестве объектов питания животного происхождения отмечены мелкие ракообразные, моллюски, насекомые, рыбы, земноводные, птенцы различных видов птиц, мышевидные грызуны. Из кормов растительного происхождения - семена риса, злаков и др. Основные способы добывания корма: склёвывание и зондирование.
Японский журавль - сильная и осторожная птица, и естественных врагов у него практически нет. Основную угрозу этим журавлям представляют антропогенные факторы. Среди них наиболее губительную роль играют осеннее и весеннее выжигание сухой травы - палы и возникающие вследствие этого пожары, которые лишают их мест пригодных для гнездования, приводят к гибели гнёзд и кладок, в результате значительная часть пар пропускает сезон размножения. Также ущерб популяции японского журавля наносят разрушение мест обитания хозяйственной деятельностью человека, браконьерство и фактор беспокойства. Также выявлена прямая связь численности японского журавля с фактором увлажнения, который складывается из суммы атмосферных осадков и условий осадконакопления. Погодные условия при этом не влияют на территориальное распределение вида в период размножения.
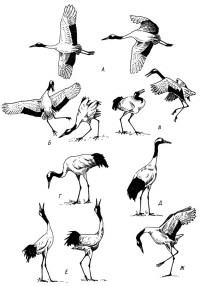 |
Рис. 4. Различные позы японского журавля.
А - журавли в полёте, Б-В - различные элементы «танцев», Г - спокойная поза во время поиска корма, Д - поза тревоги, Е - унисональный дуэт, Ж - поза агрессии.
Даурский журавль (Grus vipio Pallas, 1811) - среднего размера журавль с размахом крыльев 2,3-2,5 м и ростом 1,3-1,4 м. В гнездовое время держатся парами очень скрытно, на пролёте и зимовках образуют большие скопления. Голос как у других журавлей, но более слабый. Полового диморфизма в окраске нет.
Гнездовой ареал охватывает северо-восточную Монголию и северный Китай, среднее Приамурье, Читинскую область, Приморский край (Приханкайскую низменность). Зимовки находятся в Японии, Китае и на Корейском полуострове.
На местах гнездования даурские журавли появляются несколько позже, чем японские. Прилетают, когда болота ещё не оттаяли, а по ночам бывают сильные морозы. Некоторое время птицы держатся в стаях, а к середине апреля, когда грунт на болотах оттаивает, занимают гнездовые территории. Места обитания даурских журавлей - это открытые ландшафты степной и лесостепной зон, где он занимает травяные болота, заболоченные луга и тростниковые заросли в широких долинах рек, приозёрных котловинах, в истоках рек и ключей. На Дальнем Востоке он заселяет обширные кочкарниковые осоково-пушицевые болота и осоково-вейниковые заболоченные луга по долинам рек и котловинам озёр, перемежающиеся ленточными лесами (релками); здесь даурский журавль обитает совместно с японским.
Половой зрелости птицы достигают в возрасте 3-4 лет. Для устройства гнезда выбирают открытые места, залитые водой участки болота, иногда выгоревшие с осени и со слабо развитым травостоем, но чаще предпочитают места с достаточно густыми зарослями. Гнездо располагается прямо в воде или на кочке и имеет вид округлой хорошо утрамбованной плоской платформы с неглубоким лотком. Основание гнезда складывается из кусочков дерновины, мха, корней осоки и пропитано водой, но лоток, выстланный прошлогодними стеблями и листьями осок, совершенно сухой. Гнёзда часто используются несколько лет подряд, и тогда хорошо видны слои, в которых сохраняются остатки скорлупы от предыдущих кладок. Откладка яиц происходит в конце апреля - начале мая; в кладке, как правило, два яйца. Насиживает кладку, в основном, самка. Выводки с пуховыми птенцами впоследствии держатся неподалёку от гнездового участка, но если их тревожат, уходят в более спокойные места.
Анализ данных, полученных из Приамурья, показывает, что не все потенциальные для размножения территориальные участки заселяются в гнездовом сезоне ежегодно, также неравномерно распределена нагрузка в многолетнем цикле. Имеются территориальные участки, которые пары заселяют в течение многих лет, как правило, они соседствуют с сельхозугодьями. На них зарегистрированы лучшие показатели естественного вопроизводства даурских журавлей. Однако, имеются участки, которые журавли используют с многолетними перерывами. К ним относятся территориальные участки, которые расположены в отдалении от нарушенной среды обитания и сельхозугодий. Сравнение заселения территориальных участков в годы с обильными осадками и в годы засухи показывает, что в засушливые годы их около 60% - это территории, расположенные в хорошо увлажнённых биотопах. Гнездовой консерватизм (верность дому) у журавлей имеется, однако это не означает, что на территориальном участке не может произойти смена журавлиных пар.
В послегнездовой период биотопы с нарушенной средой обитания для журавлей предпочтительнее, чем другие. Журавли используют влажные биотопы только в гнездовой период. В биотопах с высоким уровнем воды птицы не встречаются. Не все кормовые стации входят в состав индивидуальных участков. В послегнездовой период семьи покидают гнездовой участок и пребираются ближе к суходольным лугам и сельхозугодиям. Начиная с августа, птицы кормятся на полях. Места отдыха и ночёвки входят в состав индивидуальных территорий пары; кормовые стации обычно находятся в стороне от индивидуальной территории. У птиц нет привязанности к крупным водоёмам и влажным лугам в летний период.
Во время исполнения унисонального дуэта птицы стоят параллельно друг другу и обратившись в противоположные стороны (см. рис. 5). Самец высоко поднимает крылья и запрокидывает сильно назад шею, так что клюв повёрнут вверх и назад. Самка держит крылья прижатыми к телу, запрокидывает голову, но меньше, чем самец. Клюв при этом направлен вертикально вверх. Унисональный дуэт исполняется в период гнездования и на зимовках. «Танцы» даурских журавлей сравнительно бедны элементами. «Танцы» демонстрируют как взрослые, так и молодые птицы.
Как и у японского журавля, у даурского естественных врагов практически нет; основные причины сокращения его численности, так же, как у японского, антропогенного происхождения, в первую очередь - это пожары, разрушение мест обитания хозяйственной деятельностью человека, браконьерство и фактор беспокойства.
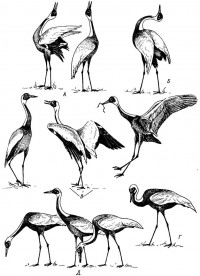 |
Рис. 5. Различные позы даурского журавля
А, Б - при унисональном дуэте; В - различные элементы «танцев»; Г - поза агрессии; Д - журавли на кормёжке
Используемая литература:
Андронов В.А. Японский (GrusjaponensisP.L.S. Müller, 1776) и даурский (GrusvipioPallas, 1811) журавли Архаринской низменности (Среднее Приамурье). Автореф. …канд. биол. наук. Владивосток, 2008. 22 с.
Глущенко Ю.Н., Нечаев В.А., Глущенко В.П. Птицы Приморского края: фауна, размещение, проблемы охраны, библиография (справочное издание) // ДВ Орн. журнал, 2010. №1, С. 3-150.
Глущенко Ю.Н., Шибнев Ю.Б., Волковская-Курдюкова. Птицы // Позвоночные животные заповедника «Ханкайский» и Приханкайской низменности. Владивосток, 2006. С. 77- 233.
Литвиненко Н.М., Нейфельдт И.А. (отв. ред.) Журавли Палеарктики (Биология, морфология, распространение). Сб. научн. тр. Владивосток, 1988. 236 с.
Панькин Н.С., Нейфельдт И.А. Даурский журавль в Амурской области // Редкие, исчезающие и малоизученные птицы СССР. Тр. Окского гос. заповедника. Вып. 13. Рязань, 1976. с. 117-120.
Флинт В.Е. Японский журавль // Птицы СССР. Курообразные, журавлеобразные. Л.: Наука, 1987. с. 280-289.
Флинт В.Е. Даурский журавль // Птицы СССР. Курообразные, журавлеобразные. Л.: Наука, 1987. с. 306-313.
Материал подготовлен к.б.н. ст.науч. сотрудником
ФГБУ «Государственный заповедник «Ханкайский»
Волковской-Курдюковой Е.А.
Использованы фотографии из архива заповедника
Наука - людям
Лотос Комарова - Nelumbo komarovii Grossh. является редким реликтовым растением, внесенным в Красную книгу РФ. Распространение в России ограничивается Приморским и Хабаровским краями и небольшой частью Приамурья. Общее распространение: Китай, п-ов Корея, Япония. Представитель древнейших цветковых, имеет научную ценность как реликт третичного периода. Описал дальневосточный лотос в 1940 году советский ботаник А.А. Гросгейм и назвал вид в честь крупнейшего ботаника академика В.Л. Комарова. Охраняется в Ханкайском (Приморский край), Хинганском (Приамурье) заповедниках.
 |
 |
Ботаническое описание. Лотос Комарова - Nelumbo komarovii Grossh. Сем. Лотосовые - Nelumbonaceae – тропическое многолетнее водное восточно-азиатское растение, стебли которого превратились в мощное толстое корневище, погруженное в подводный грунт. Одни листья у лотоса подводные, чешуевидные, другие надводные — сначала плавающие, затем приподнятые над поверхностью воды, щитовидные пластинки которых от 20 до 100 см в диаметре. Черешки длинные (до 2м длины), толстые, прямостоячие, шершавые от многочисленных шиповатых выростов. Цвет листьев сизо-зеленый от воскового налета, за счет которого они не смачиваются водой — капли воды перекатываются на них, как шарики ртути. Цветки обоеполые, надводные, одиночные до 23 см в поперечнике. В утренние часы цветок раскрывается, а к вечеру закрывается. Окраска цветка меняется в течение всего периода цветения (которое длится 2-4 дня) от ярко-розового, почти красного, до розового или бледно-розового. Они обладают слабым, но очень приятным ароматом. Чашелистиков 5-9. Лепестки многочисленные, 7-9 см длиной, 3-7 см шириной. Тычинки также многочисленные, пыльники желтые, до 2см длиной. Плод апокарпный (многоорешек). Односемянные орешки созревают в ячейках разросшегося цветоложа. Цветоложе в форме опрокинутого конуса, губчатое, содержит 20-35 погруженных темно-серых, около 1,5см длины, семян.
Фенология. Цветет лотос в конце июля – начале августа, цветение отдельных особей можно наблюдать до середины сентября. На ночь лотос закрывает цветок и опять раскрывает его утром. Цветки обладают гелиотропизмом, т.е. венчик лотоса постоянно обращен к солнцу, как бы следуя за ним. Плодоношение – сентябрь, октябрь.
 |
Местообитание. Экология лотоса довольно узкая: он любит хорошо прогреваемые мелководья в заводях и старицах, илистое дно и глубину воды не более 2м.
 |
 |
Культивирование. Лотос – декоративное водное растение. В Китае, Корее и Японии давно занимаются разведением лотоса. Культивируется это растение в странах Западной и Восточной Европы.
Размножение. Лотос хорошо размножается семенами. Однако, зрелые семена, пролежавшие некоторое время вне воды, не прорастают без предварительной обработки. Чтобы ускорить пробуждение зародышей, кожуру семян (орешков) нужно скарифицировать – надпилить со стороны конца с ямочкой. Надпиленные орешки можно вначале проращивать в комнатных условиях, затем полученные проростки высадить в открытый водоем. Семядольная часть проростка осторожно погружается в размягченный грунт на глубину 6-8 см. При этом черешки листьев оказываются почти погруженными в грунт, а листочки остаются на поверхности. Глубина воды не должна падать ниже 20 см при самом низком уровне и превышать 150 см в период половодья. Посадку лотоса под открытым небом следует производить как только исчезнет опасность поздних весенних заморозков. Для прорастания семян лотоса благоприятна температура воды, близкая к 30º С на глубине 0,5 м. Цветение у растений семенного происхождения наблюдается на 2 – 3 год жизни. Для искусственного вегетативного размножения достаточно посадить часть корневища с вегетативной почкой.
 |
 |
 |
Биологическая активность. В наше время в лотосе были обнаружены самые различные биологически активные вещества, в основном алкалоиды и флавоноиды.
Показания к применению. Применение лотоса как лекарственного растения очень многогранно. В китайской медицине корневище – тонизирующее, возбуждающее, общеукрепляющее и антитоксическое средство. На Кавказе его применяют при кишечном расстройстве, на ДВ – при астме, пневмонии и как противоядие при укусах змей. Семена стимулируют деятельность сердца. Вообще лотос входит более чем в 200 медикаментозных средств.
Примечание. Лотос - это чемпион по многим качествам: прежде всего по красоте, потом – по популярности у древних народностей. Сельское население Китая, Индии и Японии использует семена и корневища, содержащих большое количество крахмала, их едят в свежем виде, жарят, варят (как овощи), маринуют, из них делают муку. Из корневищ нередко варят суп или готовят их в качестве гарнира, подобно картофелю, а семена-орешки, жарят как каштаны. Они маслянистые, мучнистые и очень питательные.
Лотос является превосходным украшением естественных и искусственных водоемов.
Поражает великая жизнестойкость этого реликта. При неблагоприятных условиях семена лотоса не прорастают, но и не погибают, а переходят в стадию анабиоза. Неблагоприятным воздействием на произрастание лотоса являются как чрезмерное высыхание водоемов, так и резкое повышение уровня воды.
Предложения. Для сохранения и восстановления зарослей лотоса Комарова в пределах региона исследователями было предложено следующее: расширение территории Ханкайского заповедника, в частности за счёт участков его охранной зоны, содержащих водоёмы - места произрастания этого вида; создание Памятников природы в местах произрастания рассматриваемого вида, расположенных за пределами охранной зоны заповедника; восстановление зарослей в местах исчезновения; искусственная посадка семян на всех водоёмах расположенных за пределами заповедника и имеющих подходящие для произрастания условия.
Растение издавна считается священным в Индии и Китае и сейчас, цветок лотоса олицетворяет целомудрие, чистоту и духовное просветление, а обычай украшать свой дом цветами лотоса и нарцисса на Новый год происходит от поверья в их способность приносить счастье.
Помните и о том, что лотос занесен в Красную книгу России, а потому сбор его в природе для любых целей запрещен!
Используемая литература:
1. Баркалов В.Ю., Харкевич С.С. Сосудистые растения Ханкайского государственного заповедника // Ботан. Журн. Т. 81, № 11. С. 104-116.
2. Баркалов В.Ю., Харкевич С.С. 1995. Сосудистые растения государственного заповедника «Ханкайский» // Труды международной научно-практической конференции «Водно-болотные угодья международного значения: оз. Ханка». г. Спасск-Дальний. 1996. С.5-29.
3. Глущенко Ю.Н. Распространение лотоса Комарова на Приханкайской низменности и проблема его охраны // Охрана редких видов сосудистых растений советского Дальнего Востока. ДВНЦ АН СССР. г. Владивосток. 1985. С. 166-170.
4. Глущенко Ю.Н. Распространение лотоса Комарова и эвриалы устрашающей в Приморском крае // Животный и растительный мир Дальнего Востока. Вып. 3. 1997. С.
5. Красная книга РСФСР. Растения. М.: Росагропромиздат. 1988. 591с.
6. Комаров В.Л. К флоре Южно-Уссурийского края // Изв. Ботан. Сада Петра Великого. Петроград. Т. 16. Вып. 1. 1916. С. 145-180.
7. Крестов П.В., Верхолат В.П. Редкие растительные сообщества Приморья и Приамурья. Владивосток: ДВОРАН. 2003. 200 с.
8. Куренцова Г.Э. Реликтовые растения Приморья. Л.: Наука. 1968. 72с.
9. Маак К.Р. Путешествие по реке Уссури. 1-2, СПб. 1861.
10. Молчанов Г. И., Молчанова Л. П., Гулько Н. М., и др. Съедобные целебные растения Кавказа: Справочник. Ростов н/Д.: Изд. Рост. ун-та, 1989.
11. Редкие и исчезающие виды флоры СССР. 2-е доп. изд. Под ред. Тахтаджаняна А. Л. Л.: ЛО Наука, 1981.
12. Снигиревская Н. С. Семейство Лотосовые (Nelumbonaceae). С.291-293. Красная книга РСФСР (Растения). М.: Росагропромиздат, 1988.
13. Сосудистые растения советского Дальнего Востока, т. 2, с. 29–30.
14. Харкевич С.С., Качура Н.Н. Редкие виды растений советского Дальнего Востока и их охрана. М.: Наука. 1981. 231с.
Материал подготовлен младшим науч. сотрудником
ФГБУ «Государственный заповедник «Ханкайский»
Мазурок Н.Н.
Наука - людям
Деревья зимой
Совершенно безжизненными кажутся нам растения зимой. А между тем и зимой, в самые трескучие морозы, жизнь не вполне покидает деревья и кустарники. Растения в это время лишь отдыхают, накапливают силы, чтобы с наступлением весны сбросить зимние оковы. «То, что мы называем сном природы, - писал С. Покровский, - есть лишь особая форма жизни, полная глубокого смысла и значения». Такая форма жизни растительных организмов называется состоянием покоя. С приходом осени все деревья, кроме хвойных, сбрасывают листья. Природа предусмотрела это неспроста, ведь все живое должно иметь свой период покоя. Сбрасывая листву, деревья защищают себя от вымерзания, что очень важно. Как вы заметили, ели, сосны и многие другие вечнозеленые растения этого семейства, круглый год радуют глаз зелеными иголками. У этих растений сокодвижение устроено немного по-другому. Колючие иголки - это те же листья видоизмененной формы. Именно благодаря такому строению они не замерзают круглый год. Наоборот, снег или тонкий слой льда помогает дереву пережить суровые морозы.
 |
В состоянии глубокого покоя у деревьев и кустарников зимой резко заторможен обмен веществ и прекращается видимый рост. Однако это не значит, что в нем полностью остановились все процессы жизнедеятельности. Некоторые из них идут и во время зимнего покоя. Например, крахмал превращается в сахара и жиры, сахара расходуются в ходе дыхания (правда, интенсивность его зимой в 200-400 раз меньше, чем летом). Происходят в это время и процессы роста, только они никак не проявляются внешне.
Способность погружаться в состояние покоя выработалась у растений в ходе эволюции - это важное приспособление к периодическому наступлению неблагоприятных внешних условий. В состояние глубокого покоя деревья и кустарники впадают очень рано — тогда, когда погода, казалось бы, еще позволяет им нормально расти. Сигналом к покою служит для них уменьшение длины светового дня. Изменение длины дня воспринимают листья, а в отсутствие их - почки растения. Когда дни становятся короче, в растениях изменяется соотношение между фитогормонами, стимулирующими и ингибирующими процессы роста. В листьях увеличивается содержание наиболее важного природного ингибитора роста - абсцизовой кислоты, которая тормозит синтез гидролитических ферментов (амилазы, протеиназы и других), необходимых для распускания почек, прорастания семян и других процессов вегетации растений. Из листьев абсцизовая кислота транспортируется в почки и «усыпляет» их. На переход растений в состояние покоя оказывает влияние и температура. В естественной обстановке понижение температуры обычно происходит как раз в то время, когда заметно укорачивается световой день.
 |
Растениям, которые зимой укрыты снегом, под его теплой шубой не так уж холодно. Но как противостоят морозам деревья и кустарники зимой, обнаженные ветви которых насквозь пронизывает холод? Почему не гибнут их почки и побеги? Устойчивость растений к низким температурам создается главным образом благодаря внутренним изменениям в клетках, и прежде всего изменениям их химического состава. Роль антифриза - вещества, которое снижает температуру замерзания растворов, находящихся в клеточных вакуолях, играют сахара: они накапливаются в клеточном соке во время подготовки растения к зиме. Важную защитную функцию выполняют сахара и в цитоплазме: они предохраняют ее белки от коагуляции (свертывания) при понижении температуры. Чем больше сахаров накопили растения в своих клетках, тем лучше они подготовлены к действию низких температур.
Хорошо известно, что если у плодовых деревьев, например яблонь, плодоношение летом было особенно обильным, то зимой они оказываются менее морозоустойчивыми. В этом случае питательные вещества расходуются в основном на формирование плодов, а про запас сахаров откладывается мало. Неудивительно, что такие деревья легче вымерзают. Плохо переносят морозы и те растения зимой, которые осенью интенсивно росли - например, вследствие длительной теплой погоды или в результате обильной подкормки азотом. Причина здесь та же: растения к зиме плохо подготовлены, их питательные вещества использовались на рост вегетативных органов (стеблей, листьев), а не откладывались в виде запасных углеводов.
Падает морозостойкость растений и весной, когда сахара начинают использоваться в процессах жизнедеятельности, превращаться в другие соединения. Поэтому так опасны для растений весенние заморозки, хотя температура при этом далеко не достигает уровня зимних морозов, которые деревья и кустарники прекрасно перенесли. Но устойчивость растений к морозам объясняется не только накоплением сахаров в их тканях. Как показали исследования известного нашего физиолога растений, члена-корреспондента АН И. И. Туманова и его учеников, формирование морозоустойчивости - сложный, ступенчатый процесс. Свойство морозоустойчивости формируется в процессе онтогенеза растения под влиянием определенных условий среды в соответствии с генотипом растения, связано с резким снижением темпов роста, переходом растения в состояние покоя. Повышение морозоустойчивости растений тесно связано с закаливанием - постепенной подготовкой растений к воздействию низких зимних температур. Закаливание - это обратимая физиологическая устойчивость к неблагоприятным воздействиям среды.
 |
Низкие температуры просто необходимы некоторым деревьям и кустарникам во время покоя: лишь после значительного охлаждения (не менее чем до 0°С на протяжении 3-4 недель) они впоследствии могут нормально возобновить свой рост. Это было известно еще в глубокой древности; например, Плиний в своей «Естественной истории» писал: «Своевременно наступающие холода весьма способствуют укреплению деревьев, которые тогда превосходно развиваются, а в противном случае, если их ласкают австры (теплые южные ветры), истощаются, и особенно в пору цветения».
Состояние зимнего покоя - период особенно интенсивной деятельности так называемой образовательной ткани, или меристемы, из которой возникают новые клетки и ткани. Как у вечнозеленых деревьев и кустарников зимой, так и у листопадных закладываются зачатки листьев в вегетативных почках и элементы цветков - в почках цветочных. Без этого предстоящий весной переход растительного организма к активной жизнедеятельности был бы попросту невозможен. Вот почему для большого числа растений, и прежде всего для всех многолетних форм, покой - обязательное условие нормального роста в период вегетации.
 |
Литература:
Кандидат биологических наук В. И. Артамонов. Химия и Жизнь №2, 1979, с. 36-39.
Материал подготовлен м.н.с. ФГБУ «Государственный заповедник «Ханкайский»
ботаником Мазурок Н.Н..
Фотографии любезно предоставлены участниками кластера «Птицы Спасска»
Лизуновой И.В., Кузнецовой В.А..
Наука - людям
В одной из публикаций Семейного клуба экологической этики>> - рассказ о спасении раненого зимняка семьёй Шаминых. Сегодняшний выпуск рубрики «Наука - людям» подготовлен специально для Натальи Юрьевны, Дмитрия и Романа Шаминых и всех, кто принял участие в спасении попавшего в беду хищника.
Зимняк, или мохноногий канюк Buteo lagopus населяет тундру и лесотундру Евразии и Северной Америки. Зимой откочёвывает до средней Европы, Кавказа, Прикаспия, Средней Азии, Северной Монголии, Кореи; в Америке - до Калифорнии, Техаса, Луизианы, Северной Каролины. В нашей средней полосе и на юге он встречается только на пролёте и зимой, отсюда и название. Имя мохноногий (по латыни lagopus - «зайцелапый») было дано за оперение до пальцев лапы, в чём его отличие от обыкновенного канюка Buteo buteo. По окраске эти два вида несколько схожи, по размерам зимняк чуть крупнее канюка.
 |
Рис. 1. Зимняк, или мохноногий канюк Buteo lagopus
 |
 |
 |
 |
Рис. 2, 3, 4, 5. Сарыч, или обыкновенный канюк Buteo buteo
В тундре зимняк гнездится на земле - по береговым обрывам и по краям оврагов, на скалах, иногда просто на более высоких буграх, в лесотундре и северной тайге на Камчатке - на деревьях, чаще всего на лиственницах. Гнёзда устраиваются из сухих веток и иногда выстилаются сухой травой, шерстью и перьями. Кладка содержит 3-5 яиц. В тундре на гнездовании не редок, но в неблагоприятные годы число гнездящихся пар чрезвычайно сокращается (в 15 раз). Летом зимняк кормится почти исключительно северными грызунами - леммингами. В годы с низким обилием этих грызунов, зимнякам приходится перемещаться по тундре в поисках кормных мест. На пролёте и зимой держится на окрытых пространствах, главным образом, в речных долинах и в культурном ландшафте, в это время большую долю в его рационе составляют полёвки. Как у всех тесно связанных в питании мышевидными грызунами хищных птиц, численность зимняков и в гнездовой области, и на пролётах и зимовках сильно меняется по годам. Это объясняется усиленным размножением в лемминговые годы, а также повышением смертности и негнездованием в годы неурожая грызунов.
Охотится зимняк или скрадом, сидя на земле и ожидая приближающуюся добычу, или на лету, летая кругами медленно и низко, около 8-10 м над землёй, и временами «трясясь» на месте, как пустельга. Эта особенность зимняка во время охоты, подобно пустельге, высматривать в воздухе добычу, трепеща на одном месте, позволяет в природе легко отличать его от близких видов. Иногда стережёт добычу, сидя у нор. Хорошо передвигается по земле. Держится обособленно и лишь в местах, изобилующих грызунами, иногда собирается группами и даже стаями. На ночлег усаживается на деревья.
Зимняк - обычный зимующий в Южном Приморье вид, являющийся самым многочисленным представителем соколообразных здесь в зимний период. Отмечается с начала октября до начала мая. Вскоре после прилёта занимает удобные для охоты участки, но часто после снегопадов численность и перераспределение зимняков меняется. Населяет обширные открытые пространства со слабо выраженным рельефом, сельскохозяйственные земли. Охотно использует различные присады, такие как деревья, кусты, кучи соломы, разнообразные конструкции (например, опоры ЛЕП) и нежилые постройки, но зимняка часто также можно увидеть сидящим на земле. Численность зимняка широко варьирует по годам, находясь в зависимости от обилия основной добычи и толщины снежного покрова. Их обилие в долине р. Раздольная составило 0,3-1,3 ос./км2, в среднем, 0,9 ос./км2 (по данным: 2002-2007 годов). В равнинной местности Ханкайско-Раздольненского водораздельного эразионного плато она менялось, в среднем, в разные годы (1997-2008) от 0,05 до 1,61 ос./км2. На Приханкайской низменности - от 0,09 до 1,1 ос./км2. По данным полученным при изучении питания зимняков, проводящих зиму в Южном Приморье, доля участия объектов составила: большой полёвки - от 51 до 85%, полевой мыши - от 5,9 до 34%, мыши-малютки - от 0,3 до 0,5%, серой крысы - от 0,5 до 0,9%, землероек - от 0,3 до 0,7%. Не часто, но довольно регулярно в зимний период в южных районах Уссурийского края¹ регистрируются тёмные особи зимняка, называемые меланистическими (или, просто, меланистами), доля таких особей в Южном Приморье составляет около 0,7% зимующей здесь популяции (n=2500; по данным за 1986-2004 годы).
¹ Уссурийский край - в Российской империи общее название местности между реками Уссури, Сунгача, озером Ханка с одной стороны и берегом Татарского пролива и Японского моря — с другой. Располагался на территории административных Уссурийского и Южно-Уссурийского округов Приморской области. В настоящее время территория Уссурийского края примерно соответствуют границам современного Приморского края и части Хабаровского края Российской Федерации.
Литература:
1. Волковская-Курдюкова Е.А. Птицы агроландшафтов Южного Приморья. Дис. … канд. биол. наук. Владивосток, 2011. 270 с.
2. Волковская-Курдюкова Е.А., Курдюков А.Б. Необычно высокая концентрация хищных птиц-мышеедов зимой 2001/2002 годов в Южном Приморье // Русский орнитологический журнал. Экспресс- выпуск, 2003. Вып. 208. С. 3-16.
3. Воробьёв К.А. Птицы Уссурийского края. М.: изд-во АН СССР, 1954. С. 75.
4. Галушин В.М. Хищные птицы леса. Жизнеописания, проблемы, решения. М.: изд-во «Лесная промышленность», 1980. С. 66-67.
5. Глущенко Ю.Н., Шибнев Ю.Б., Волковская-Курдюкова Е.А. Птицы // Позвоночные животные заповедника «Ханкайский» и Приханкайской низменности. Владивосток, 2006. С. 116-117.
6. Дементьев Г.П. Отряд Хищные птицы. Мохноногий канюк, или зимняк // Птицы Советского Союза. Т. 1. М.: Гост. изд-во «Совет. наука», 1951. С. 307-314.
7. Корелов М.Н. Отряд Хищные птицы Falconiformes. Зимняк// Птицы Казахстана. Т. II. Алма-Ата: изд-во АН Каз. ССР, 1962. C. 685-688.
8. Назаренко А.А., Глущенко Ю.Н. Меланистические особи зимняка Buteo lagopus как маркёры североамериканской популяции Buteo lagopus sanctijohannis, зимующей в южных районах Уссурийского края // Русский орнитологический журнал. Экспресс-выпуск, 2005. Т.14. Вып. 299. С. 833-835.
9. Семаго Л. Аристократы неба. Рассказы, очерки. Воронеж: Центрально-Черноземное книжное изд-во, 1981. 222 с.
10. Штегман Б.К. Фауна СССР. Птицы. Дневные хищники. Т. 1. Вып. 5. М.-Л.: изд-во АН СССР, 1937. С. 212-217.
Материал подготовлен к.б.н. ст.науч. сотрудником
ФГБУ «Государственный заповедник «Ханкайский»
Волковской-Курдюковой Е.А.
Фотографии любезно предоставлены
научным сотрудником БПИ ДВО РАН
к.б.н. Курдюковым А.Б.